郑重声明:原文参见标题,如有侵权,请联系作者,将会撤销发布

FRONTIERS IN NEUROROBOTICS, (2017)
Abstract
行走的动物在运动过程中根据它们的情况产生适应性的肢体间协调。肢体间协调是通过神经系统、肌肉骨骼系统和环境的动态相互作用产生的,尽管其潜在机制尚不清楚。最近,对生物适应机制的研究引起了人们的关注,基于关于感觉运动相互作用的神经生理学发现的仿生控制系统正在为腿式机器人开发。在这篇综述中,我们介绍了由各种因素(运动速度、环境情况、身体特性和任务)引起的腿式机器人的自适应肢间协调。此外,我们展示了自适应肢间协调的特征属性,例如步态滞后和不同的时间尺度适应。我们还讨论了实现自适应肢间协调的潜在机制和控制策略以及腿式机器人控制系统的设计原理。
Keywords: legged robot, interlimb coordination, adaptation, sensorimotor interaction, central pattern generator
1. INTRODUCTION
动物通过巧妙地操纵其复杂和冗余的肌肉骨骼系统来产生适应性运动行为。运动是日常生活中需要的一种重要行为。根据情况选择步态,如速度和环境,是一种突出的适应性运动功能。人类双足行走并使用步行和跑步步态。四足动物使用四只腿,产生行走、小跑和疾驰的步态。六足昆虫使用六只腿,创造异时(波浪)、四足和三足步态以及形成连续体的中间步进模式。这些步态,包括六足动物的过渡和中间步进模式,是通过腿部运动的肢内和肢间协调产生的。肢内协调是一条腿内节段或关节之间的关系,而肢间协调是腿之间的关系。例如,在肢体内协调的自适应控制中,踝关节跖屈、膝关节伸展和髋关节伸展的峰值时间在人类步行步态中是异相的,但在人类跑步步态中它们是移动的并且几乎同相(Diedrich et al., 1998)。在肢体间的自适应控制中协调,腿之间的脚步顺序发生变化(Muybridge, 1957),该顺序主要用腿运动之间的相对相位来解释,因为腿运动是周期性的,每条腿的周期几乎相同(请注意,由于踏步模式的灵活性,有时会在昆虫中观察到腿之间的不同频率,Pearson and Franklin, 1984)。在四足行走步态中,虽然左右腿反相运动,但同侧前后腿没有。相反,在四足小跑步态中,同侧前后腿以及左右腿反相运动;也就是说,对角线腿同相移动(Hildebrand, 1965)。为阐明步态机制而进行的测量数据分析表明,步态的选择是基于代谢和生物力学因素(Margaria, 1938; Hoyt and Taylor, 1981; Farley and Taylor, 1991)。然而,关于这些因素在决定步态中的作用的报告(Hreljac, 1993; Minetti et al., 1994; Raynor et al., 2002; Wickler et al., 2003)是相互矛盾的,因此潜在的机制仍不清楚。
为了阐明动物的适应性运动功能,已经独立进行了神经生理学和生物力学研究。神经生理学研究主要研究神经系统的结构和活动,而生物力学研究通常研究肌肉骨骼系统的功能作用。然而,运动是通过神经系统、肌肉骨骼系统和环境之间的动态相互作用产生的。因此,很难从单一的角度全面分析运动机制。此外,步态被视为此类复杂动力系统中的自组织模式(Schöner et al., 1990; Diedrich et al., 1998; Griffin et al., 2004; Schilling et al., 2013a)。步态的稳定性结构已经从扰动的响应中识别出来,特别是通过相位振荡器和相位响应曲线(Couzin-Fuchs et al., 2015; Funato et al., 2016)基于相位减少理论(Kuramoto, 1984)。然而,由于运动中动力因素之间相互作用的复杂性,很难理解稳定性结构是如何产生的。为了充分阐明运动机制,需要对神经和肌肉骨骼系统进行综合研究,以找到产生适应性运动行为的过程。
最近,为了揭示运动机制,腿式机器人引起了人们的关注。带有执行器(如电动机、气动和液压执行器)的机器人机械系统已被用于研究肌肉骨骼系统在运动中的动态作用。机器人的控制系统是根据神经生理学的发现开发的,并采用了各种传感器,如触摸传感器、称重传感器、加速度传感器、陀螺仪传感器、激光测距扫描仪和视觉系统。这种方法使我们能够通过神经系统、肌肉骨骼系统和环境之间的动态相互作用来模拟和研究步态生成。特别是,位于脊椎动物脊髓和无脊椎动物胸神经节中的中枢模式发生器(CPG)是阐明运动机制的重要因素(Grillner, 1975; Orlovsky et al., 1999; MacKay-Lyons, 2002)并帮助开发了腿式机器人的运动控制系统。CPG是一组相互连接的神经元,可以在不需要感觉反馈的情况下被激活以产生运动模式。支持这一假设的证据最初是由Brown (1911)提出的。除了开环控制功能外,CPG还接收感官反馈以调节运动指令。这种感官反馈的闭环结构对于根据情况实现自适应行为至关重要。已经通过使用神经网络或振荡器网络提出了各种CPG模型,并在控制腿机器人中实现[参见Ijspeert (2008)的评论]。例如,Taga和Shimizu (1991)以及Taga (1995)对人类双足运动的CPG模型进行了开创性研究。他们为身体机械模型采用了铰接式多连杆系统,并为CPG模型采用了由Matsuoka (1985)开发的神经振荡器。该CPG模型接收用于运动的局部和全局信息的感觉信号。他们证明适应性运动是通过身体动力学、振荡器动力学和环境之间的相互作用建立的。他们称之为"全球夹带"。尽管可以通过纯粹的自反控制机制(Cruse et al., 1998; Manoonpong et al., 2007; Lewinger and Quinn, 2011; Schilling et al., 2013a,b)和经典机器学习控制(Bongard et al., 2006; Cully et al., 2015)而不是使用CPG模型,CPG概念和建模对腿式机器人的研究产生了很大的影响。
在这篇综述中,我们关注运动中肢体间协调的自适应控制。我们引入了由各种因素(运动速度、环境状况、身体特性和任务)引起的动物和腿式机器人的自适应肢间协调。此外,我们还展示了动物和机器人适应性肢体协调的特征,例如步态滞后和不同的时间尺度适应。最后,我们讨论了实现自适应肢体间协调的潜在机制和控制策略以及腿式机器人控制系统的设计原理。

2. ADAPTIVE INTERLIMB COORDINATION IN ANIMALS AND ROBOTS
2.1. Speed-Dependent Adaptation
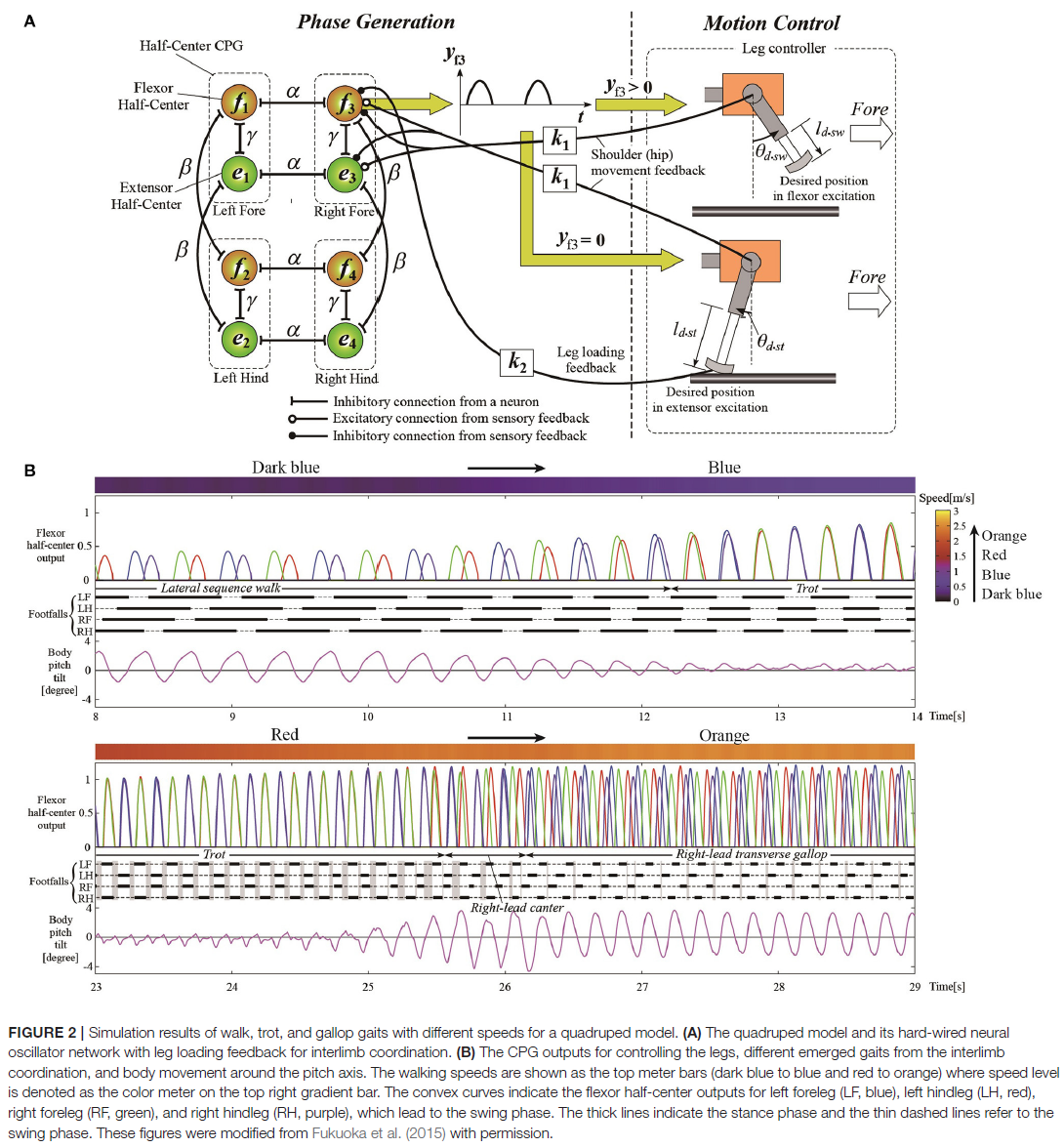
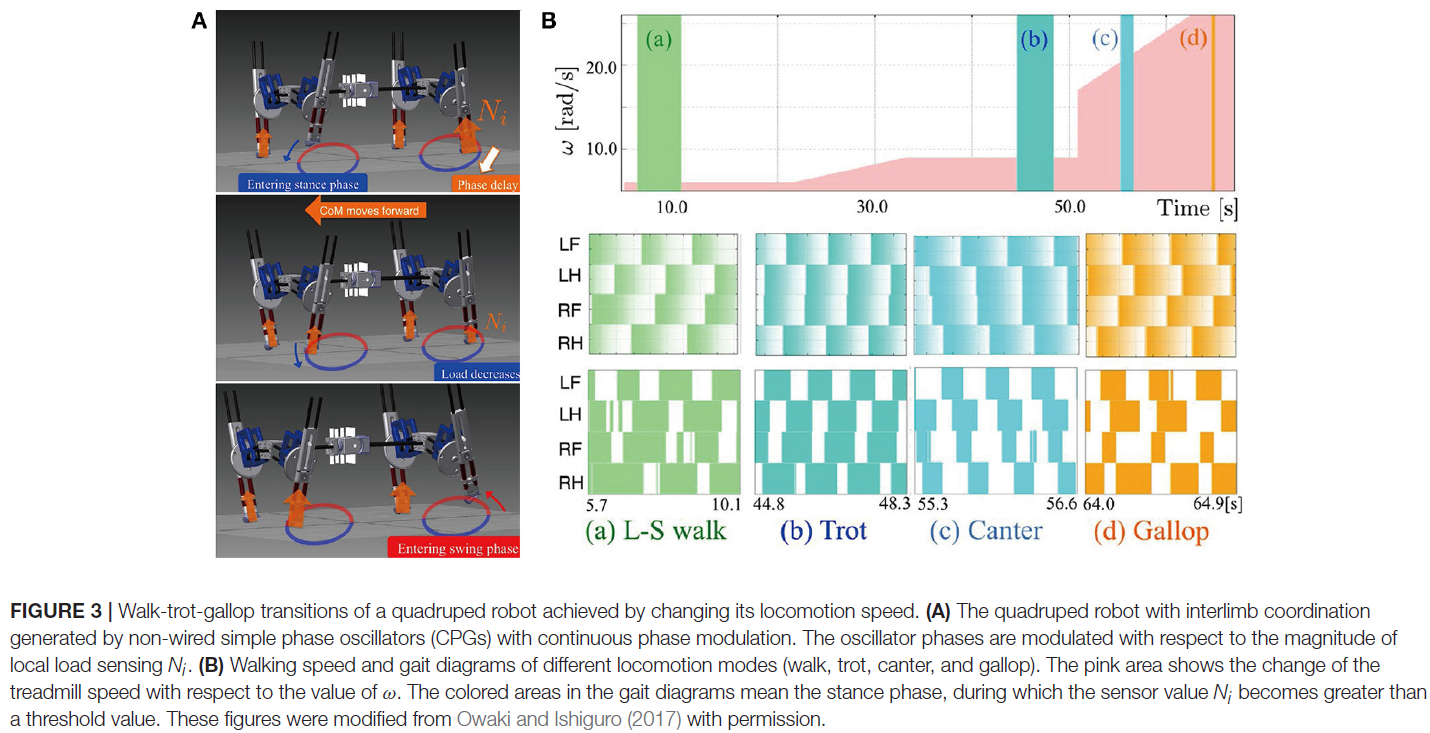
当改变有腿动物的运动速度时,会出现最普遍的适应性肢体间协调。甚至在跑步机上的脊椎猫中也观察到了这一点(Forssberg and Grillner, 1973; Orlovsky et al., 1999),其中腿之间的相位关系发生变化,步态在步行、小跑和疾驰之间发生变化。在报告的研究中,猫的脊髓是从大脑中切下的,但它们仍然通过脚和腰带之间的接触获得感官反馈。感觉信号随着皮带速度的变化而变化,从而引起他们的步态转变。这一结果突出了感觉运动相互作用对适应性肢体协调的重要贡献。四足机器人通过对具有局部感觉反馈的脊柱CPG进行建模,实现了依赖于运动速度的自适应肢间协调(Maufroy et al., 2010; Aoi et al., 2011, 2013b; Owaki et al., 2013; Fukuoka et al., 2015; Owaki and Ishiguro, 2017)。这可以在以下示例中看到。图1显示了Aoi et al. (2013b)的四足机器人、控制系统和步行-小跑过渡的实验结果(该机器人在步态转换中表现出滞后,如第3.1节所述)。图2,来自Fukuoka et al. (2015)的工作,展示了四足动物的步态从慢速步行过渡到中速小跑,以及高速横向疾驰。图3来自Owaki和Ishiguro (2017)的工作,还显示了四足机器人在没有神经耦合的情况下,从横向序列(L-S)行走到小跑甚至到疾驰的自发步态转变。这些机器人研究使用简单的神经振荡器或相位振荡器用于CPG模型,并从振荡器相位产生腿部运动。更具体地说,一个振荡器产生了一条腿的运动,振荡器之间的相位关系决定了步态。每个振荡器相位都通过腿部的局部感觉信息进行调节,例如足部接触和腿部负荷,仅发生在一条腿上。
作为这些机器人研究中的重要控制架构,振荡器之间的相位关系没有预定义,振荡器只是弱耦合或解耦。也就是说,步态不是由使用强耦合的振荡器动力学决定的(Schöner et al., 1990; Canavier et al., 1997; Ito et al., 1998; Golubitsky et al., 1999),而是由通过局部感觉反馈的全身动力学和振荡器动力学。肢体间协调仅以神经动力学、身体动力学和环境之间的自组织方式产生。
根据步态速度的类似适应性肢体间协调也出现在六足昆虫中,例如竹节虫(Wilson, 1966; Graham, 1972; Crus, 1990; Grabowska et al., 2012)、蟑螂(Hughes, 1952; Delcomyn, 1971; Pearson, 1976; Bender et al., 2011)和苍蝇(Strauß and Heisenberg, 1990; Wosnitza et al., 2013; Berendes et al., 2016)。特别是,竹节虫和苍蝇会根据步态速度平滑地改变它们的肢体间协调(Wilson, 1966; Graham, 1972; Wosnitza et al., 2013)。更具体地说,腿之间的相对相位对于步态速度以线性方式连续变化。这类似于一些哺乳动物,包括绵羊,但不同于其他哺乳动物,包括狗。在诸如狗之类的哺乳动物中,步态转换具有相对的腿部相位,这些相位以sigmoid方式突然变化(Alexander and Jayes, 1983)。尽管有人认为蟑螂主要通过CPG本身实现肢体间协调(Fuchs et al., 2011),但CPG本身并不会产生用于竹节虫行走的协调运动模式,因为感觉反馈很重要(Bässler and Wegner, 1983; Büschges et al., 1995; Büschges et al., 2008)。Cruse和他的同事提出了一种名为Walknet的人工神经网络,它根据六种不同的规则控制腿部运动,通过感官信息来调节肢体间的协调(注意单条腿的控制器在没有CPG的情况下运行)。这些规则是从竹节虫的行为实验凭经验得出的[参见Cruse et al. (1998), Dürr et al. (2004)和Schilling et al. (2013a)的评论]。其中三个规则是通过在光滑的表面上干扰腿部运动而设计的。规则根据相邻腿的感官信息改变了一条腿的循环持续时间。由于感觉运动相互作用,Walknet控制的昆虫模型产生了连续的运动模式,如三脚架、四脚架和波浪步态,以及在竹节虫中观察到的中间步进模式。此外,这些模型还用于各种情况,例如在不平坦的表面上行走(Kindermann, 2002)、腿部截肢(Schilling et al., 2007)、谈判曲线(Schilling et al., 2013b)以及爬过大间隙(Bläsing, 2006),其运动行为与竹节虫相当。Tóth和Daun-Gruhn (2016)基于Hodgkin-Huxley动力学开发了神经网络模型,并将其与肌肉骨骼模型相结合,以解释昆虫的肢体间协调机制。尽管他们的模型没有产生在竹节虫(Wilson, 1966; Graham, 1972)和果蝇(Wosnitza et al., 2013)中观察到的中间步进模式,他们的结果表明,不同腿的提肌-下肢神经肌肉系统之间的联系对于复制三足和四足步态的主要特征是必要的。Ambe et al. (2013, 2015)以类似于上述四足机器人的方式,为六足机器人使用具有足部接触信息的局部感官反馈的简单相位振荡器。他们通过具体的感觉运动交互产生了连续的运动模式,例如异时和三脚架步态以及中间步进模式,而不是根据运动速度预先定义模式。此外,显示的一个重要方面是竹节虫站立期间的正速度反馈(Bässler, 1976)。正速度反馈已在机器人上进行了测试(Schmitz et al., 2008)。
类似地,蜈蚣等多足动物会根据步态速度改变它们的四肢协调。多足类动物的身体轴长而灵活,当步态速度增加时会产生身体起伏(Manton, 1965)。除了起伏的幅度增加外,同侧腿部运动的相位关系随着起伏的体节运动而变化。Aoi et al. (2007, 2013a)开发了一种具有六个体节和十二条腿的多腿机器人,它使用扭转弹簧来实现身体轴的灵活性。机器人通过增加运动速度,通过直线行走的超临界Hopf分叉表现出身体起伏,因此表现出身体起伏对速度的依赖性,类似于蜈蚣表现出的依赖性。

2.2. Environment-Dependent Adaptation
在动物和机器的移动运动中使用腿的优点是通过操纵脚接触位置,即使在复杂的环境中也能获得高可穿越性。但是,腿式机器人的可穿越性还远没有达到动物的水平。在运动过程中,腿部运动包括站立阶段(脚与地面接触)和摆动阶段(脚离开地面)。在站立阶段,腿部支撑身体抵抗重力,并通过脚和地面的相互作用产生推进力和减速力来移动身体。地面的几何特性各不相同。这些属性包括平坦地形、倾斜地形或不规则和崎岖地形。地面的物理性质也会发生变化。这些性质包括坚硬而滑的地面如石头,柔软的地面如松散的土壤,以及流动和渗透的地面如沙子。脚和地面之间的相互作用对于产生运动至关重要,需要根据地面情况实时适应运动行为。动物实际上根据环境情况表现出适应性的肢体间协调。要控制有腿的机器人,阐明和应用动物的动力学原理至关重要。
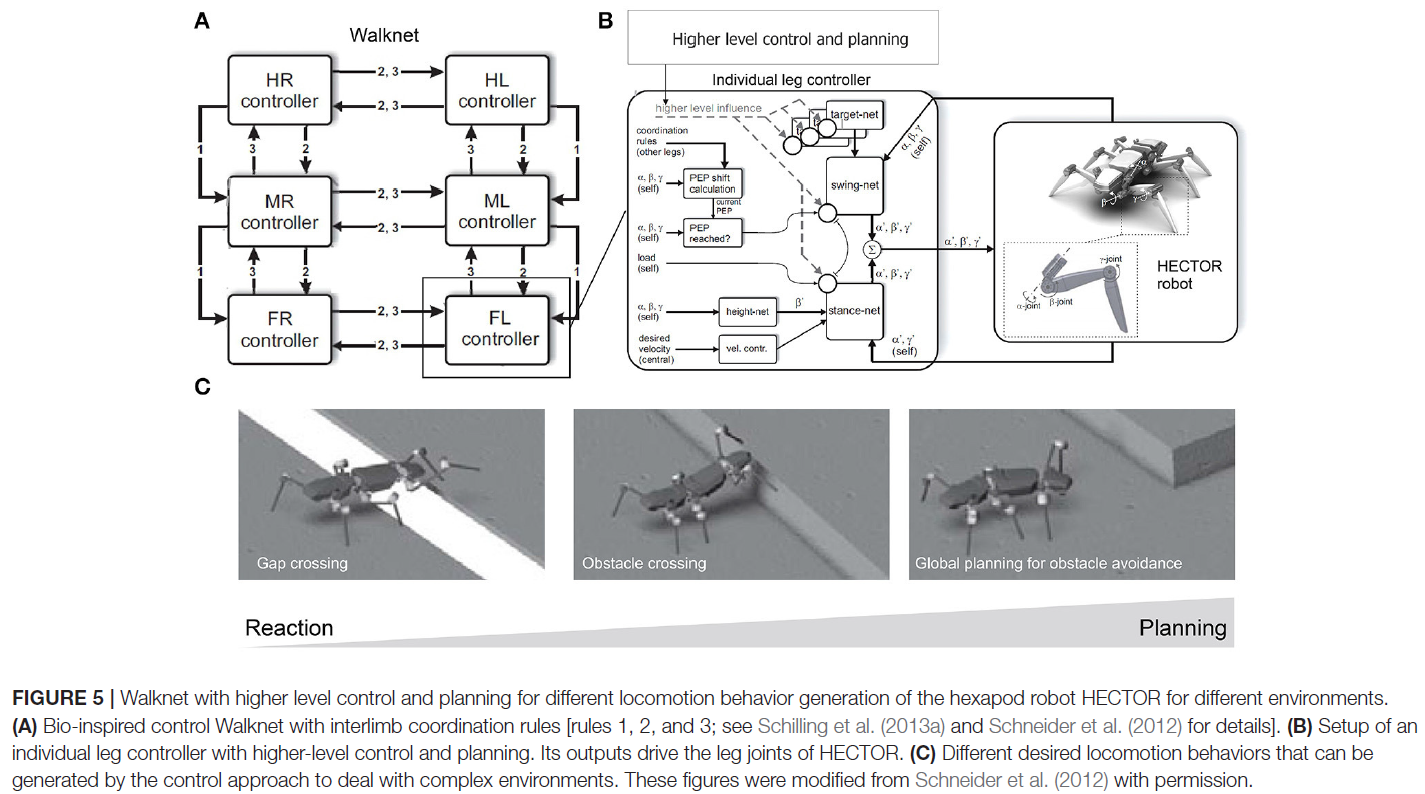
Manoonpong和他的同事为腿式机器人开发了一系列基于神经CPG的模块化运动控制(Manoonpong et al., 2008, 2013; Steingrube et al., 2010; Goldschmidt et al., 2014; Xiong et al., 2014, 2015; Dasgupta et al., 2015; Grinke et al., 2015)。他们表明,使用这种控制方法可以实现自适应肢间协调,使机器人能够应对复杂环境,例如在困难地形上行走(Steingrube et al., 2010; Manoonpong et al., 2013; Goldschmidt et al., 2014; Xiong et al., 2014, 2015; Dasgupta et al., 2015)并在未知的杂乱区域避开障碍物(Manoonpong et al., 2008; Grinke et al., 2015),如在昆虫中观察到的。例如,他们使用基于混沌CPG的自适应网络和六足机器人的感官反馈实现了模块化神经控制(图4A, B; Steingrube et al., 2010)。由于CPG的内在混沌动力学类似于在某些生物CPG中观察到的动力学(Rabinovich and Abarbanel, 1998),因此利用动力学来根据环境条件生成各种步行模式。在他们的设置中,机器人展示了标准行走的四足步态,上坡行走的波浪步态,下坡行走的波浪和四足混合步态,以及快速行走的三脚架步态以执行快速趋光性(图4C)。然而,这种离散步态的实现并不一定对应于在昆虫中发现的情况。除了这些多种步态之外,混沌动力学尤其有助于从地面上的洞中自行解开一条腿(图4B),从而增强了立足点搜索行为。在Dasgupta et al. (2015), Goldschmidt et al. (2014)和Manoonpong et al. (2013)将前向模型集成到模块化神经控制中,使机器人能够有效地预测其行走状态,以便在复杂地形上行走时在摆动和站立阶段伸展或抬高其腿。通过这种设置,机器人使用四足步态在不平坦的地形上行走,并使用波浪步态爬过高障碍物以及爬上一段楼梯。此外,它通过使用毛毛虫步态成功跨越了一个大间隙,其中每对左腿和右腿同时移动。然而,在这种情况下,竹节虫表现出比毛虫协调更复杂的行为,毛虫协调很少被采用(如果有的话)(Blaesing and Cruse, 2004)。在Xiong et al. (2014),通过引入基于虚拟激动剂-拮抗剂机制(VAAM)的肌肉模型扩展了模块化神经控制,并产生神经机械控制以实现腿部顺应性。将神经机械控制与感觉运动学习相结合,可以使用具有相应腿部顺应性的不同步态实现节能步行(Xiong et al., 2015)。机器人有效地在不同的表面上行走,包括海绵、砾石、细砾石和草地。为了适应在杂乱环境中避开障碍物,将具有突触可塑性的自适应神经感觉处理网络引入模块化神经控制(Grinke et al., 2015)。自适应处理网络可以通过短期机器人记忆驱动不同的转向行为。因此,机器人四处走动并调整其转弯行为,以避开不同情况下的障碍物并避免出现尖角或死锁(图4D)。除了模块化神经控制方法外,Schneider et al. (2012)开发了仿生控制,它将Walknet(上面提到的)与更高级别的控制和规划(图5A, B)相结合,用于六足机器人HECTOR的自适应肢间协调。通过使用这种控制技术,可以生成多种行为(例如,穿越间隙、穿越障碍物以及避免或攻击障碍物的全局规划)来处理复杂的环境(图5C)。此外,Schilling和Cruse (2017)扩展了Walknet以发明新行为并在实际使用它们之前通过内部模拟对其进行测试。Arena et al. (2017)提出了基于CPG的多层运动控制与昆虫启发的运动技能学习。它可以自适应地协调类似果蝇的六足机器人的四肢,以实现稳定的行走和越障。
当马走上斜坡时(Wickler et al., 2003)或当它们负重时(Farley and Taylor, 1991),小跑到疾驰的过渡速度会降低。竹节虫、蟑螂和甲虫等六足昆虫会根据地面坡度改变步态(Spirito and Mushrush, 1979; Pelletier and Caissie, 2001; Grabowska et al., 2012)。此外,虽然蟑螂在正常行走时使用三脚架步态,但当它们被拴在支撑球上以减少负载时,步态会改变为异时态(Spirito and Mushrush, 1979);上坡和负重会对他们的步态产生类似的影响(Tang and Macmillan, 1986)。Fujiki et al. (2013a)扩展了用于六足步行者的四足机器人(图1B)的控制系统,并表明步态在三脚架和异时步态之间通过感觉运动相互作用而改变,这取决于负载和倾斜角,如在昆虫中观察到的。
Fukuoka et al. (2003), Fukuoka and Kimura(2009)以及Kimura et al. (2007a, b)使用Matsuoka (1985)开发的神经振荡器来控制四足机器人(Tekken系列)。他们根据感觉信息整合了各种反射模型,例如屈肌反射、伸肌反射和前庭脊髓反射。此外,他们模拟了强直迷宫响应以调整滚动运动以与俯仰运动同步。机器人在不规则地形(例如台阶和斜坡)上产生稳健的运动,同时诱导步行和小跑之间的步态转换。
当地面像沙子一样流动时,腿在运动过程中会深入地面。因此,与地面相互作用以产生升力、阻力和推力变得复杂[见Aguilar et al. (2016)的评论]。Li et al. (2009, 2013)为六足机器人使用三脚架步态,并通过使用机器人在颗粒介质中移动的力模型调整腿部形状和腿部运动,产生类似于硬地的运动性能。
除了对斜坡、崎岖地形、杂乱区域和可流动区域的适应之外,还研究了处理非对称环境条件的肢体间适应。对于不对称情况,分带式跑步机已用于人类研究(Dietz et al., 1994; Reisman et al., 2005; Morton and Bastian, 2006)、猫(Yanagihara and Kondo, 1996; Frigon et al., 2013)、小龙虾(Müller and Cruse, 1991a, b)和竹节虫(Bässler and Wegner, 1983; Foth and Graham, 1983)。跑步机有两条平行的带独立控制的速度,因此能够人为地创造左右对称和不对称的步行环境(并列配置:皮带之间的速度相同,分带配置:皮带之间的速度不同)。虽然细节将在3.2节中讨论,但根据皮带速度条件已经观察到自适应肢间协调。这种适应甚至出现在脊椎猫身上(Forssberg et al., 1980; Frigon et al., 2013)。Otoda et al. (2009)为二维双足机器人开发了一种没有CPG模型的感觉驱动控制器,Fujiki et al. (2013b)将简单的相位振荡器用于双足机器人的CPG模型,该模型具有足部接触信息的局部感官反馈,与上述实现自适应肢体间协调的四足和六足机器人类似(Aoi et al., 2011, 2013b; Ambe et al., 2013, 2015; Fujiki et al., 2013a)。双足机器人在分离式跑步机上实现了自适应肢间协调。


2.3. Body-Dependent Adaptation

2.4. Task-Dependent Adaptation
动物经常遇到必须改变运动行为的情况。例如,当行走路径上出现障碍物时,他们越过障碍物,或向右或向左转以避免与障碍物发生碰撞(这也与环境适应有关)。这样的任务主要是通过调节腿部运动产生的,因此肢体协调的自适应控制很重要。然而,同样重要的是肢体间协调的自适应控制。要越过障碍物,前肢首先清除障碍物,然后后肢跟随它。前肢的脚必须比平时抬得更高,以避免与障碍物发生碰撞,这种运动会延迟脚的接触。尤其是双足和四足动物,前肢足部接触后必须抬起后肢足;否则,避障任务将失败,因为对侧肢体在抬起后肢时不支撑身体(Aoi et al., 2013c)。
改变步行方向的转向行为用于各种任务,例如目标追踪(Szczecinski et al., 出版中)和避障(图4D, 5C)。Knops et al. (2013)通过使用基于Hodgkin-Huxley动力学的神经网络模型控制了竹节虫中间腿的机械模型,并通过在光滑表面上行走的竹节虫观察到的两种不同策略产生转向行为:内中腿由前向侧转,或由前向后踏步。在Aoi et al. (2016)中,蜈蚣状多腿机器人的转弯机动性通过改变身体轴柔度而由Hopf分叉引起的直线行走不稳定性得到增强。尽管腿伸直的节肢动物的重心低,因此不能有效地倾斜,但腿直立的哺乳动物的重心高,可以利用身体倾斜来帮助转动。由于转动运动的左右不对称性,人体转动中腿之间的相对相位从反相转变(Courtine and Schieppati, 2003)。在Aoi和Tsuchiya (2007)中,具有关于足部接触信息的局部感官反馈的简单相位振荡器被用于双足机器人的转弯行走,就像在分带式跑步机上行走一样。腿之间的相对相位根据转弯半径移动,以补偿身体倾斜引起的左右不对称;这种转变使机器人能够实现高转弯性能。
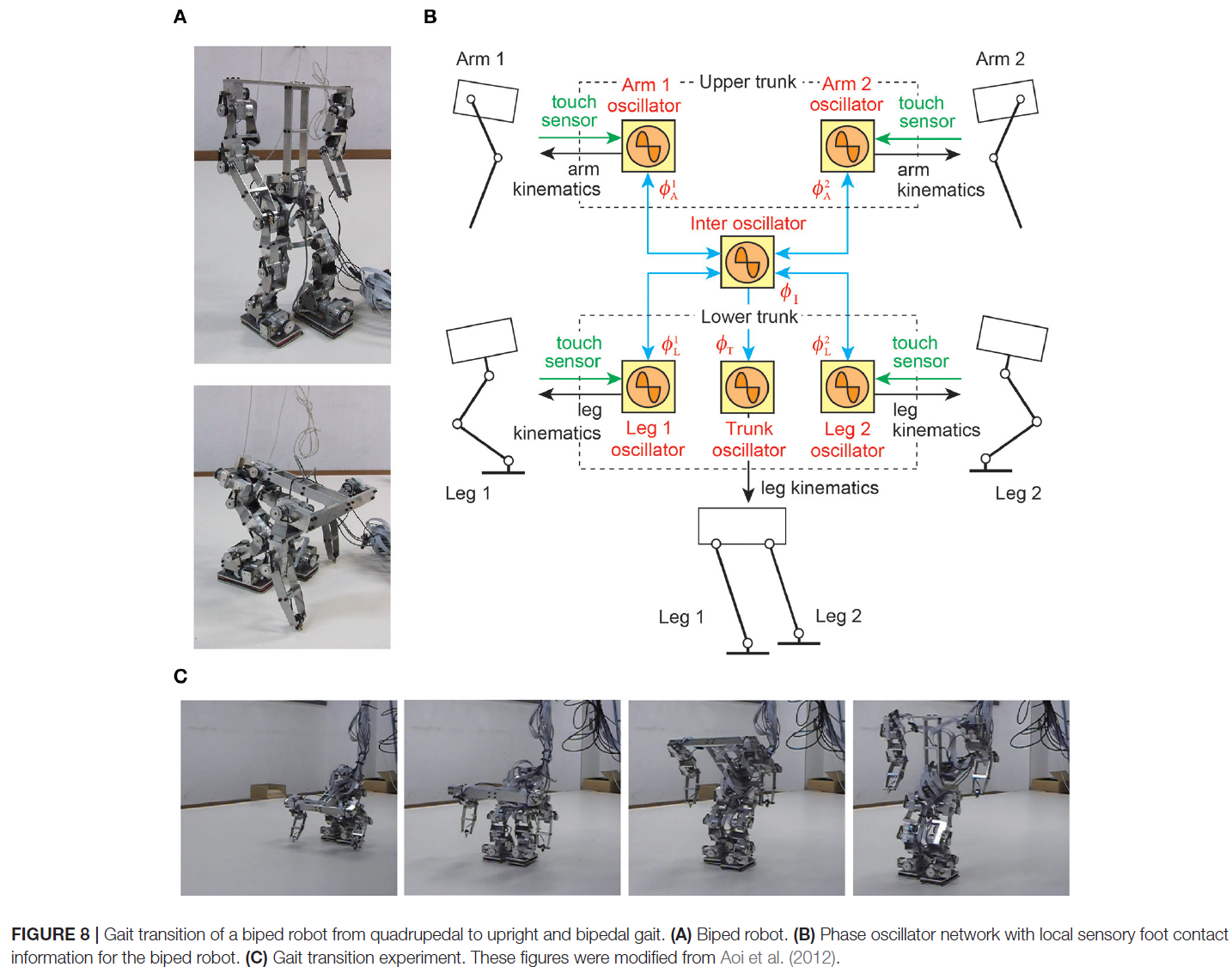
从四足步态到直立和双足步态的转变对于腿式机器人来说是一项具有挑战性的任务,因为它需要剧烈的运动变化(Asa et al., 2009; Aoi et al., 2012; Kobayashi et al., 2015)。特别是,由于机器人必须抬起躯干以使手臂离开地面,因此支撑肢体位置和质心位置之间的适当关系很重要。也就是说,足够的肢体间和躯干协调是至关重要的;否则,机器人很容易摔倒。在Aoi et al. (2012)中,通过手臂和腿的地面接触信息进行感官调节的简单相位振荡器用于双足机器人(图8A, B)。该控制器基于运动学协同的概念进行了扩展(Freitas et al., 2006; Ivanenko et al., 2007; Latash, 2008; Funato et al., 2010)以改变机器人的步态转换运动,并允许机器人成功地将步态从四足变为直立和双足(图8C)。
腿式机器人可用于搜索和救援任务。在这种情况下,地面不仅是不规则的,而且是脆弱的,就像一个碎片散落和建筑物倒塌的区域,当受到外力(例如机器人腿的压力)时,该区域的表面可能会倒塌。在这种情况下,重要的是通过使用腿部的触觉信息来检查地面状况,以确保稳定行走。在Ambe和Matsuno (2016)中,提出了一种用于地形确定的具有触觉感官反馈的控制机制。通过控制机制,四足机器人在将腿放在地面上时,可以通过其力传感器感知立足点是否稳定。此外,这种机制可以产生足够的肢体间协调,因此即使在探针运动中立足点倒塌,机器人也不会绊倒。因此,机器人可以有效地在不稳定的地形上行走,避免绊倒和造成周围区域的大面积坍塌(图9)。还提出了其他方法来基于触觉反馈和图像信息估计脆弱和滑溜的立足点(Tokuda et al., 2003; Hoepflinger et al., 2010, 2013)。
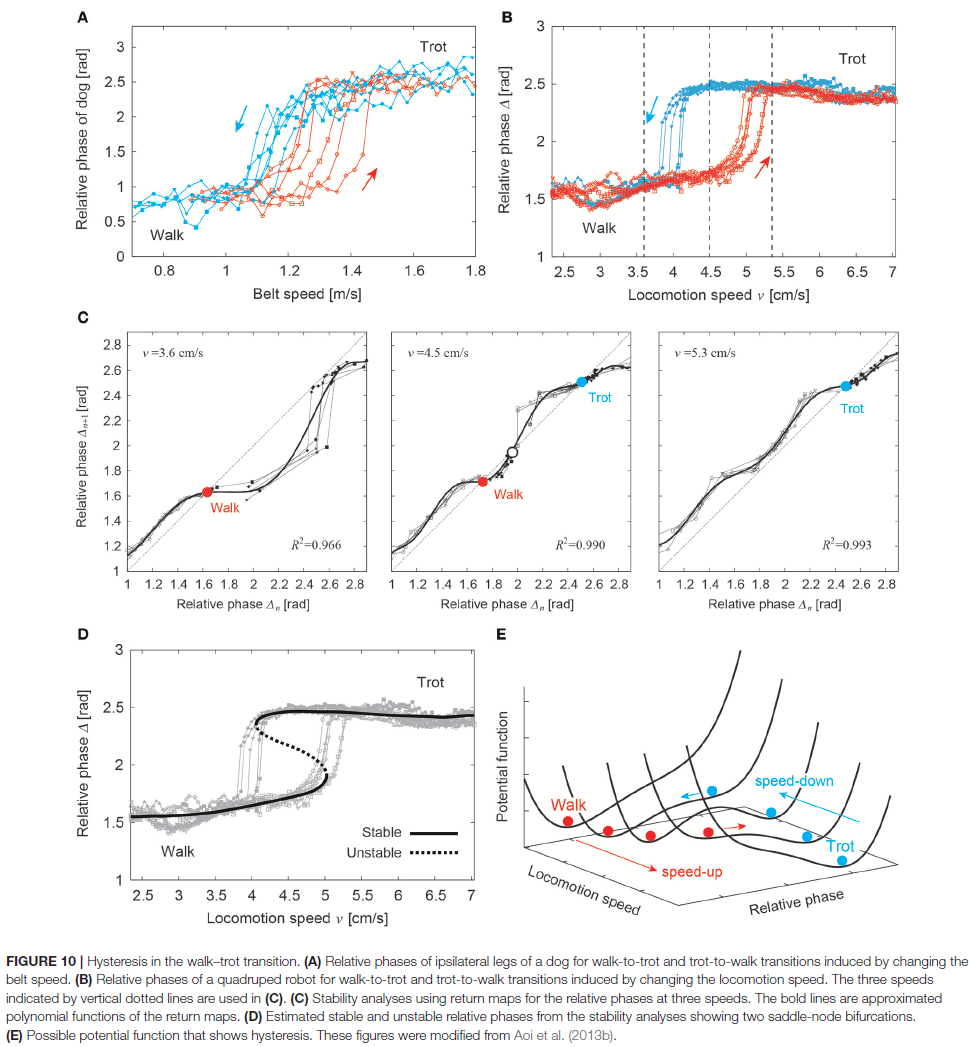
3. CHARACTERISTIC PROPERTIES OF ADAPTIVE INTERLIMB COORDINATION
3.1. Hysteresis in Gait Transition
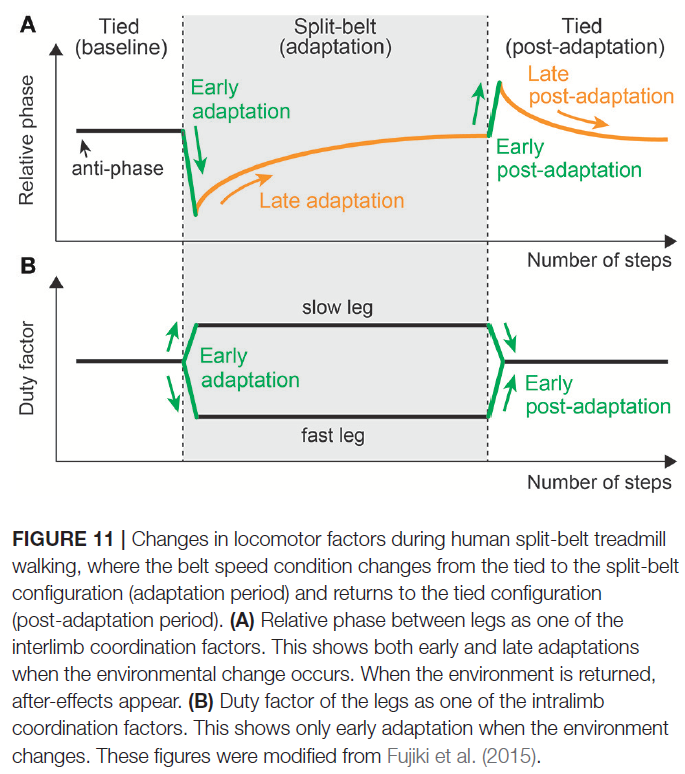

3.2. Two Different Time-Scale Adaptations
当环境突然变化时,运动行为会迅速调节以适应环境变化,并逐渐显示出逐渐调节以获得新的运动模式。这种行为表明发生了运动学习。这已在运动过程中的肢体间协调中观察到。尤其是上面提到的分带式跑步机行走就是一个很好的例子。
(未完待续)
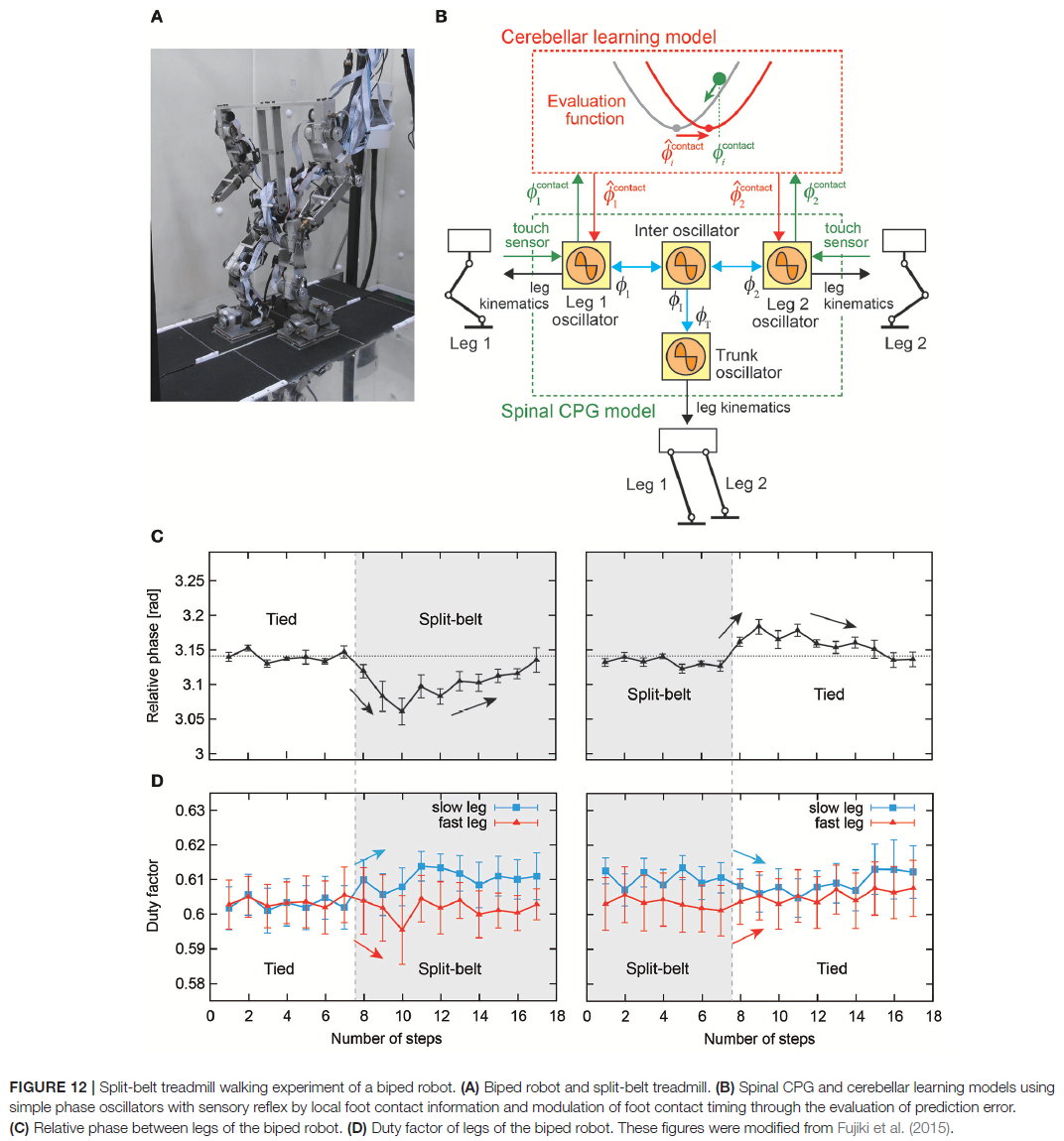
4. KEY FACTORS AND MECHANISMS FOR ADAPTIVE INTERLIMB COORDINATION
在前面的部分中,我们介绍了动物和腿式机器人的自适应肢间协调,以处理不同的运动速度、环境情况、身体特性和任务。在此,我们讨论了肢体间协调自适应控制的关键因素和机制。
关键机制之一是CPG,它位于脊椎动物的脊髓和无脊椎动物的胸神经节中。除了拮抗兴奋性运动神经元的反相活性外,毛果芸香碱诱导的节律特征似乎与在竹节虫中观察到的任何运动输出相对应(Büschges et al., 1995)。然而,神经生理学研究表明,CPG对运动很重要(Grillner, 1975; Orlovsky et al., 1999; MacKay-Lyons, 2002)。CPG是一组相互连接的神经元,可以在不需要感觉反馈的情况下被激活以产生运动模式。如Ijspeert (2008)所述,从使用Hodgkin-Huxley神经元的详细生物物理模型(Traven et al., 1993; Cataldo et al., 2006; Holmes et al., 2006; Bungay and Campbell, 2009)提出了具有不同复杂程度的各种CPG模型;以及使用LI神经元或IF神经元的连接主义模型(Buchanan, 1992; Arena, 2000)使用耦合振荡器抽象模型(Ijspeert et al., 2007; Chung and Slotine, 2010; Yu et al., 2014)。尽管一些机器人研究表明,复杂的昆虫行为,例如连续的步态转换、在包括大间隙的不规则地面上行走,以及以不规则步幅的曲线行走,可以在没有CPG模型的情况下进行复制(Cruse 等人,1998 年;Lewinger 和 Quinn,2011;Schilling 等人,2013a,b),这些 CPG 模型改进了腿式机器人的运动控制,例如速度控制 (Ijspeert, 2008) 和对感官噪声和感官故障的鲁棒性(Di Canio et al., 2016)。特别是控制腿式机器人的关键问题是前馈和反馈控制器的设计以及这些控制器的集成。CPG模型为我们的设计和集成提供了有用的想法,以便集成控制器以生物学上合理的方式工作(有和没有CPG模型的控制器之间的比较将有助于发现CPG模型的贡献)。
大多数研究采用了抽象的CPG模型,这些模型与运动单元有硬连线,用于生成不同的基本运动行为,例如步行和游泳。可以通过使用简单的外部输入信号在不同的步态或运动模式之间切换(Kirchner et al., 2002; Ijspeert et al., 2007; Manoonpong et al., 2008)。尽管作为开环控制的CPG是产生基本规律运动的关键,但感觉反馈是适应不同速度、环境、身体和任务所需的一个非常重要的因素,如前面的自适应肢体间协调部分所述。将CPG与感官反馈相结合,可实现具有适应性的闭环控制。对于机器人实施,影响CPG活动的不同感觉反馈包括本体感觉反馈(例如,关节/腿部运动和力)和外感觉反馈(例如,足部接触和视觉)。这种反馈可以调节CPG活动的频率、相位和幅度[参见Buschmann et al. (2015)的评论]。
频率调节(也称为夹带,Buchli et al., 2006)使用反馈信息来调整CPG的频率,以便反馈和CPG的频率同步(Nachstedt et al., 2017)。通常,关节角度反馈用于机器人研究中的这个过程(Endo et al., 2004; Buchli and Ijspeert, 2008; Di Canio et al., 2016),频率调制主要用于适应运动速度(Harischandra et al., 2004, 2011; Di Canio et al., 2016)和身体变化(Ren et al., 2015)。相比之下,相位调节通常使用足部接触和足部负荷反馈来调整CPG的相位,以根据情况调节摆动和站立阶段的持续时间。特别是,经常用于腿式机器人相位调节的相位重置机制是根据猫的触觉传感器反馈的相移和规则重置行为发展而来的(Conway et al., 1987; Duysens, 1997; Schomburg et al., 1998; Rybak et al., 2006; Frigon et al., 2010)和竹节虫(Büschges, 1995; Bässler and Büschges, 1998)。已经通过与肌肉骨骼模型和肌肉协同假设的整合研究了相位重置的功能作用(Aoi et al., 2010, 2013c; Aoi and Funato, 2016),并且控制策略在腿式机器人中实现并有助于改善(Tsuchiya et al., 2002; Aoi and Tsuchiya, 2005, 2007; Nomura et al., 2009; Aoi et al., 2011, 2012, 2013b; Ambe et al., 2013, Fujiki et al., 2015 ., 2013a, b, 2015)。相位调节也被广泛用于不同的适应,包括运动速度(Tsuchiya et al., 2002; Aoi et al., 2011, 2013b; Ambe et al., 2013, 2015; Fujiki et al., 2013a; Owaki et al., 2013; Fukuoka et al., 2015; Owaki and Ishiguro, 2017), 环境条件(Aoi and Tsuchiya, 2005; Aoi et al., 2010; Fujiki et al., 2013a, b, 2015),身体特性(Tsuchiya et al., 2002; Aoi et al., 2011; Fujiki et al., 2013a; Owaki et al., 2013; Barikhan et al., 2014)和任务(Aoi and Tsuchiya, 2007; Aoi et al., 2012, 2013c)。幅度调节使用不同类型的反馈(例如力和视觉)来调节CPG的幅度。这种调节是通过运动前神经元网络间接实现的(Buschmann et al., 2015)。Goldschmidt et al. (2014)和Grinke et al. (2015)通过使用视觉反馈进行环境依赖适应来采用这种策略,例如六足机器人爬过障碍物或避开障碍物。
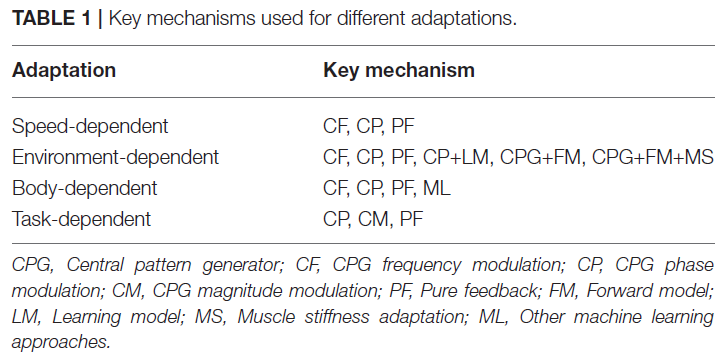
还可以通过将这些CPG调节技术与其他仿生方法(例如自适应肌肉刚度控制)相结合来实现自适应肢间协调(Xiong et al., 2015)。Manoonpong et al. (2013)表明,将运动命令或情感副本转化为预期感官反馈的仿生前向模型是环境依赖性适应的重要组成部分,即在不同的地形上行走。通过使用分体式跑步机,Fujiki et al. (2015)表明,小脑学习模型在最小化预测误差的同时调节运动命令对于环境依赖性适应也很重要。表1粗略地对用于不同适应的关键机制进行了分类。
除了这些受生物启发的关键因素(CPG、感觉反馈、前向模型、学习模型和肌肉僵硬),它们通常应用于独立控制单个腿或关节,大多数研究明确设计完整的肢间连接以获得 所需的运动行为。这导致了适应性和灵活的肢体间协调的局限性(例如,Kirchner et al., 2002; Ijspeert et al., 2007; Harischandra et al., 2011; Manoonpong et al., 2013; Ren et al., 2015)。为了克服这些限制,提出的替代范式通过局部传感、身体-环境相互作用和弱耦合或解耦CPG实现了肢体间协调(Tsuchiya et al., 2002; Aoi et al., 2011, 2013b; Shim and Husbands, 2012; Ambe et al., 2013, 2015; Fujiki et al., 2013a; Owaki et al., 2013; Barikhan et al., 2014; Owaki and Ishiguro, 2017),而不是通过预定义的肢间连接。尽管所提出的范式在肢体间协调方面具有很高的灵活性和适应性,但它有时会遇到不稳定的运动。如上所述,基于感觉反射调节CPG相位的相位重置是在保持运动稳定性的同时获得灵活和自适应的肢体间协调的解决方案之一(Tsuchiya et al., 2002; Aoi and Tsuchiya, 2007; Aoi et al., 2011, 2012, 2013b, c; Ambe et al., 2013, 2015; Fujiki et al., 2013a, b, 2015)。然而,这仅使用相位调节并且具有局限性。因此,该方向的一项未来研究是找到一种可以自主形成塑性连接的方法,以实现稳定但仍然灵活和自适应的运动。此外,CPG、感觉反馈、身体动力学、前向模型、学习模型和肌肉僵硬的相互作用,以实现高度适应性、鲁棒性和节能的运动仍有待探索。
5. CONCLUSION
尽管步行动物通过巧妙地操纵其复杂和冗余的肌肉骨骼系统来创造适应性运动行为,但其潜在机制仍不清楚。为腿式机器人设计控制架构以自主实现这种适应性仍然是一个挑战。尽管一些腿式机器人在没有生物系统启发的情况下通过纯粹的工程方法产生了自适应运动行为,但神经生理学的发现,如CPG组织和感觉运动相互作用,对于设计腿式机器人的控制系统很有用。从动态角度来看,使用CPG模型和感官反馈的机器人实验对于以神经动力学、身体动力学和环境之间的自组织方式理解步态生成和适应是很有见地的。在这篇综述中,我们展示了由各种因素(如运动速度、环境状况、身体特性和任务)引起的动物和腿式机器人运动中的自适应肢间协调。我们还展示了适应性肢体间协调的特征,例如步态滞后和不同的时间尺度适应。腿式机器人正在成为了解包括肢体间协调在内的运动机制的宝贵工具。未来,随着机器人系统(例如执行器和传感器)的改进,通过与神经和肌肉骨骼系统的复杂模型(例如Hodgkin-Huxley模型和肌肉肌腱单元模型)的集成来增强生物学的合理性和可行性将变得非常重要,并通过与简单模型(例如模板模型)集成来提取动态特征(Full and Koditschek, 1999; Holmes et al., 2006)。此外,进一步改进和发展分析方法也很重要,例如减相理论(Kuramoto, 1984)和协同分析(Ivanenko et al., 2004; Latash, 2008),从多重和冗余数据中阐明基本因素。