郑重声明:原文参见标题,如有侵权,请联系作者,将会撤销发布!

Cell 2020
Summary
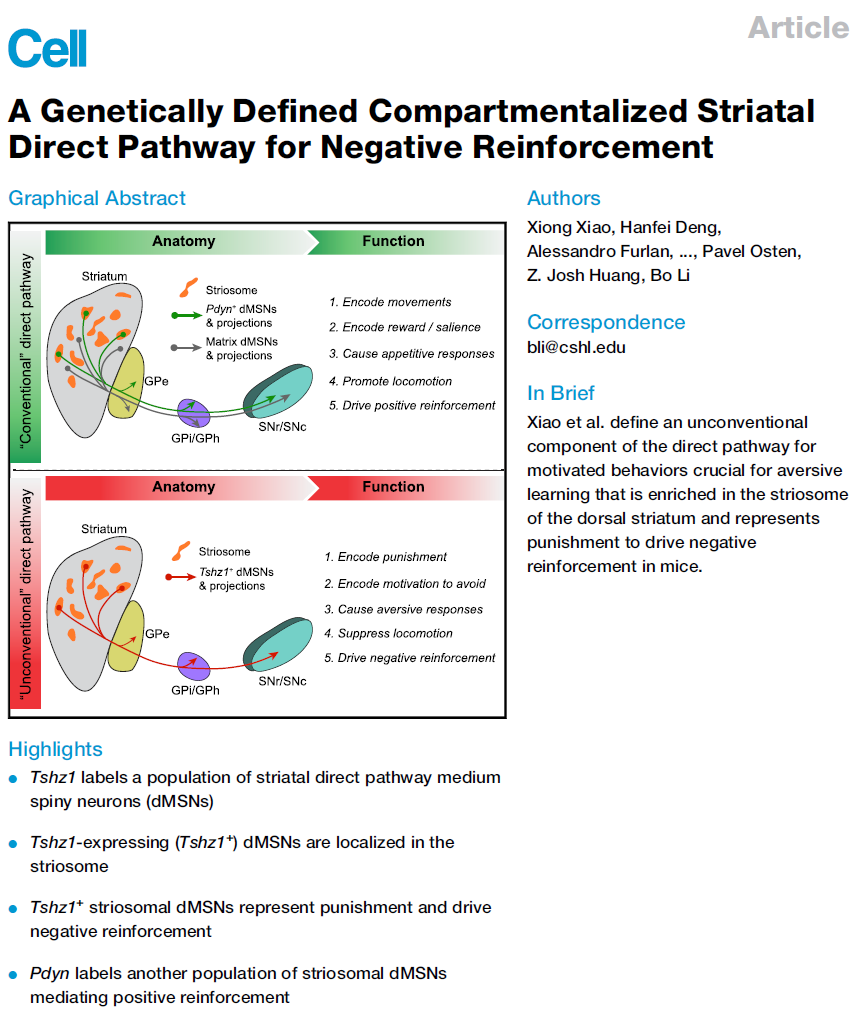
背侧纹状体内的纹状小体区室与强化学习和动机调节有关,但是纹状体神经元如何促进这些功能尚不清楚。在这里,我们显示了遗传确认的纹状体群体,该群体表达茶衫族锌指1 (Tshz1)并属于直接途径,驱动负强化,对于小鼠厌恶性学习至关重要。与"常规"纹状体直接途径相反,Tshz1神经元一旦被激活,就会引起厌恶,运动抑制和负强化,它们会收到一组独特的突触输入。这些神经元主要是受惩罚而不是奖励而兴奋,它们代表了对惩罚的预期或逃避的动机。此外,抑制这些神经元会损害基于惩罚的学习,而不会影响奖励学习或动作。这些结果建立了纹状体神经元在通过惩罚强化的行为中的主要作用,此外还揭示了经典模型中未说明的直接途径的功能。
INTRODUCTION
背侧纹状体(DS)具有多种行为功能,包括运动控制,强化学习和动机调节。它主要由表达D1型或D2型多巴胺受体的中棘神经元(MSN)组成,分别引起所谓"直接途径"和"间接途径"。一种流行的模型认为,直接和间接途径具有相反的功能,前者促进运动并促进奖励或正强化,后者抑制运动并促进厌恶或负强化。然而,最近的研究表明,运动过程中直接和间接途径神经元被共激活,而不具有拮抗动作。这些发现导致针对运动控制的两种途径的模型进行了修订,并且更加细化或更详细。然而,在正和负的强化中这些途径的二分观点保持不变。
支持DS在强化学习中作用的主要证据来自体内记录研究,表明DS神经元具有结果评估所需的价值信息。同时,广泛的实验工作和计算模型表明,纹状小体是一种在DS中嵌入周围基质的神经化学独特区室,它起着评估作用,并在强化学习中扮演"critic"的角色。此外,纹状小体特别容易受到情绪和动机障碍的影响,表明这种纹状体区室可能在调节情感和动机过程中也起着重要作用。因此,这些发现表明,纹状小体调解或保留了DS的主要功能。
然而,尽管进行了深入研究,但迄今为止,尚未明确定义纹状小体中神经元的功能。特别是,纹状体神经元如何促进强化学习或调节动机尚不清楚。研究纹状体神经元的一个主要挑战在于这样一个事实,即该质体呈迷宫状,并且没有清晰的解剖边界,这使得很难用目前可用的方法进行有针对性的体内记录或操作。为了解决这个问题,在这项研究中,我们利用小鼠遗传学靶向特定的纹状体神经元群体。这一策略为我们发现"非常规"纹状体直接途径奠定了基础,该途径来自纹状小体中富含的D1神经元的子集,并且在激励行为中具有意想不到的作用。
RESULTS
Genetic Targeting of Neurons in the Striosome
Tshz1+ dMSNs and Pdyn+ dMSNs Have Opposing Behavioral Functions
Tshz1+ dMSNs Are Preferentially Excited by Aversive Stimuli
Valence Representations and Valence-Specific Predictive Signals in Tshz1+ dMSNs
Tshz1+ dMSNs Represent Distinct Aspects of Avoidance Behavior
Tshz1+ dMSNs Are Required for Aversive Learning
DISCUSSION
Tshz1+ dMSNs Are "Unconventional"
我们的结果表明,Tshz1在DS中划分了一个独特的MSN群体,纹状小体充满该群体,并构成了"非常规"直接途径,对负强化和厌恶性学习至关重要。Tshz1+ dMSN的功能以及这些神经元对运动的抑制作用与先前描述的"常规"直接途径的功能相反。相比之下,Pdyn+ dMSNs(纹状小体充满该群体)显示出类似于常规直接途径的特性,促进了奖励或正强化并促进了运动。Tshz1+ dMSN和Pdyn+ dMSN之间的功能差异可能是由于这些神经元的不同连通性引起的。实际上,我们表明这两个种群收到不同的单突触输入集。
这些神经元还可以差异地投射到突触后目标神经元上并调节其功能。在这方面,最近的研究表明,纹状小体MSN是向habenula-projecting GPi (GPh)神经元输入的主要来源,这对于评估强化学习期间的负和正结果至关重要。因此,可以想象的是,Tshz1+ dMSN和Pdyn+ dMSN分别通过激活(通过抑制)或抑制GPh神经元传达有关惩罚和奖励的信息,从而有助于评估行为结果或激发价特定的行为(图S7L)。作为替代或补充,Tshz1+ dMSN和Pdyn+ dMSN可能通过投射到中脑多巴胺区域(图S7L)上来影响学习或动机,该区域已显示可接收来自纹状体神经元的直接输入。这两个群体如何精确调控GPh神经元或多巴胺神经元以影响行为,这是未来研究的重要问题。未来的研究还应该评估Tshz1+ dMSN和Pdyn+ dMSN之间的连通性,并阐明这两个群体在行为过程中如何相互作用。
Tshz1+ dMSNs Contribute to Distinct Aspects of Negative Reinforcement
最近的影像学研究表明,分别针对D1-Cre或A2A-Cre转基因小鼠的DS中单独dMSN或iMSN的活动与动物自发的自然运动的速度相关。对多巴胺能给DS的投入也有类似的观察。这种相关性可能代表了基质神经元的功能,由于基质区室占DS的比例要比基质小得多,因此可以优先成像。我们发现在主动回避任务中,许多Tshz1+ dMSN的活动似乎也与动物的奔跑速度有关。但是,与跑步相关的Tshz1+ dMSN并不能简单地代表此任务的运动,因为这些神经元的主要群体("失败细胞")仅在失败试验中由喘气引起的反应性跑步期间被激活,但在成功的试验中,不会因为期待喘气而主动跑步,而另一群体("成功细胞")则表现出相反的行为。
重要的是要认识到,主动回避任务中的运动速度可以反映动物的情感状态,而这种情感状态在简单的自发运动中可能不会发生变化。因此,在反应性奔跑期间的小鼠速度可以反映出惩罚的感知值(即气喘),而在主动奔跑过程中的速度可以反映未来惩罚的预期价值,因此可以避免攻击的活力。在这种情况下,Tshz1+ dMSN激活与反应性或主动跑步之间的表面关系实际上可能表明这些神经元参与了不同的情感过程。当动物遭受惩罚时,失效单元可以编码惩罚的价值,并有助于评估和更新负结果。这一概念与以下观点一致:Tshz1+ dMSN编码气喘的价值(图4F和4G;图S4F和S4G),并且在执行/不执行任务时厌恶性学习需要Tshz1+ dMSN的活动(图7;图S7)。另一方面,成功细胞可以表示未来惩罚的预期价值,因此有助于在预期惩罚时避免回避。此功能类似于先前描述的"负动机"或"平均显著性"。ND细胞可能在评估和振奋功能中都发挥作用,因为它们在体验和预期喘气时都会被激活。尽管抑制Tshz1+ dMSN不会影响学习后的no-go反应(图7),但这可能反映了以下事实:在训练有素的动物中,"no-go"(即,no-licking)反应需要很少的努力。是否需要Tshz1+ dMSN活动才能激发更严格的主动回避。
Tshz1+ dMSN的发放也可以通过刺激的感觉属性来调节,例如气喘和冲击的感觉属性。尽管如此,与运动速度一样,这些神经元的反应本身并不仅仅代表感觉属性,因为,例如,成功细胞对气喘没有反应,而ND细胞却与气喘无关地被激活。总体而言,我们的结果表明,Tshz1+ dMSN的编码可以最好地描述为针对厌恶性刺激以及厌恶性刺激引起或预期的行为动作和情感状态。
总而言之,尽管巴甫洛夫条件反射任务使我们能够在操作上将"负价神经元"识别为Tshz1+ dMSN的主要群体,但基于主动回避任务的结果却发现,这些神经元可以归类为亚群,即"失败细胞","成功细胞"和"ND细胞"——在强化学习中可能扮演不同的角色。
Toward Understanding MSN Diversity and Striosome Function