1.1 生命之树
1、生物学家的梦想
一句话:构建生命之树。
2、方法与途径
最理想的途径是通过化石证据,然而化石之零散和不完整,因此大多数研究者转为比较形态学和比较生理学。通过这两个途径,基本可以得出有机体进化历史的主题框架。但是,形态及生理性状进化之复杂,不可能产生一副清晰的图像。
相比比较形态学和比较生理学,分子生物学的优势:
1)DNA只有4种碱基。因此可以用来比较所有有机体的进化关系,这是前2个途径不能做到的
2)DNA进化演变多少是有规律的,因而能用数学模型来描述其变化,并可用来比较进化关系较远的生物间的DNA
1.2 进化机制
进化原理:
进化的首要原因:基因突变。核苷酸插入/缺失、替代、重组和基因转换等引发的突变基因或者DNA序列,通过群体水平的遗传漂变或/和自然选择,最终在物种中得以固定。如果该基因产生新的形态或者生理 性状,除非基因在此突变,否则次性状会传递给所有后裔。因此,对某一类群构建有效的系统树,我们就可以找到具有此突变性状的谱系。
应用:
1)上述信息可以用来研究特定性状的进化机制。将具有某特定性状的谱系所处的环境条件与无此性状的谱系所处的环境条件进行比较,就可能会搞清楚该性状是由自然选择还是随机演化所决定的。如果我们能鉴定出所涉及的基因并研究其进化演变,将会明了何种类型的突变产生了特定的形态或生理性状。
2)研究突变、自然选择、遗传突变和重组等的相对重要性是群体遗传的一个重要的课题。对一个基因座上的不同等位基因测序,来了解他们的进化历史。这里问题不是不同物种间的系统树,而是同一个物种内不同等位基因的系统树。
3)多态等位基因的系统发育分析可以为研究两个群体间的基因交流的程度提供重要信息。
1.3 基因的结构和功能
从功能来看,基因分为蛋白编码基因和RNA编码基因。RNA编码基因产生tRNA、rRNA、snRNA 等,这些非信使RNA是RNA基因编码的最终产物。结构如下图:
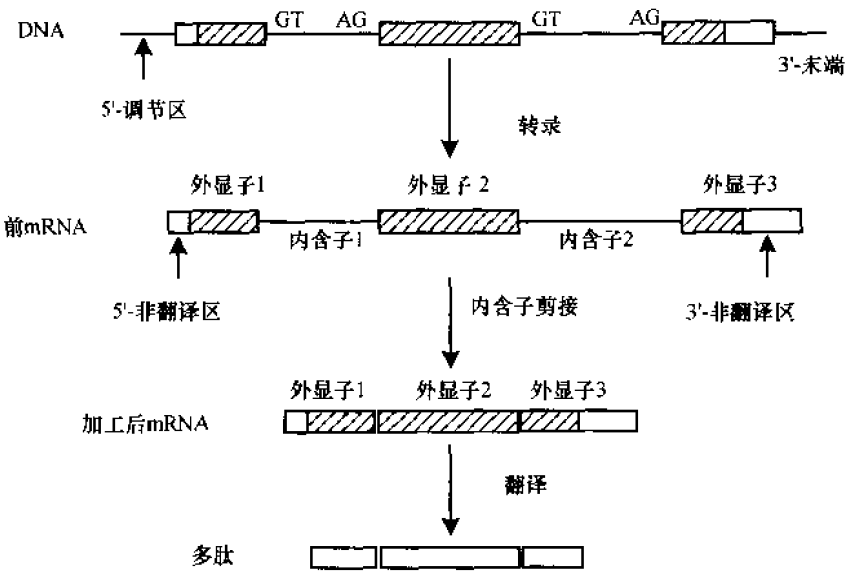
大多共用一套密码子。在研究DNA序列进化的时候,起始密码子是必须要排除掉的,因为他们是多数处于不变的状态。同理,终止密码子也应该去掉。
1.4 DNA序列的突变
DNA序列的突变基本类型:核苷酸插入、缺失、倒位、替代(颠换、转换)。

因为遗传密码的特性,大多数同义替代出现在第三位,少量出现在第一位,第二位不是非同义就是无义突变。
短的插入缺失显然是由于复制差错,长的插入缺失借助于不等交换或者DNA转座,基因插入的另一种可能是物种间的水平基因组转移,显然也算是转座因子介导的。不等交换在DNA含量增减的进化过程中发挥重要作用,特别是免疫球蛋白和核糖体RNA基因等这样的多基因家族中,不等交换对产生基因的多拷贝有重要的作用。与不等交换相关的另一个遗传事件是基因转换。基因转换会改变一个DNA片段,使其与另一个片段完全相同,该事件是由异源DNA错配修复引起。它具有使基因家族成员同质化的能力,但不改变基因的拷贝数。
1.5 密码子的使用频率
密码子使用偏倚的原因:
1、同功tRNA 的相对丰度
高表达基因的密码子使用频率与其细胞内的同功tRNA的丰度相关。翻译机器倾向于使用更丰富的同功tRNA来产生蛋白,特定tRNA的丰度同基因组中编码此tRNA 的基因拷贝数相关。
适度中等表达的基因,编码同一氨基酸的所有密码子有平均使用的倾向。或许是由于翻译速度不快,因而稀有的同功tRNA也能被用到缘故。
核苷酸突变多少是随机的,但是在高表达基因中,与丰富的tRNA不匹配者,将被净化选择所淘汰。在中度表达基因中,选择压如此之低,以致于各种同义密码子都可以使用。
2、偏倚的突变压
例如细菌中G+C的相对频率的变化范围在25%-75%之间,主要是由于核苷酸水平上GC至AT的正向和回复突变压的差异造成的。
为维持蛋白功能,由突变压决定的平衡频率会同预期的密码子第三位的GC含量不同,因为某些密码子第三位的替代导致氨基酸的替代而受到净化选择的淘汰作用;密码子第二位,全是非同义,所以该替代主要是功能制约而不是突变压制约;密码子第一位由于一小部分是同义替代,因而突变压介于第二位和第三位之间。
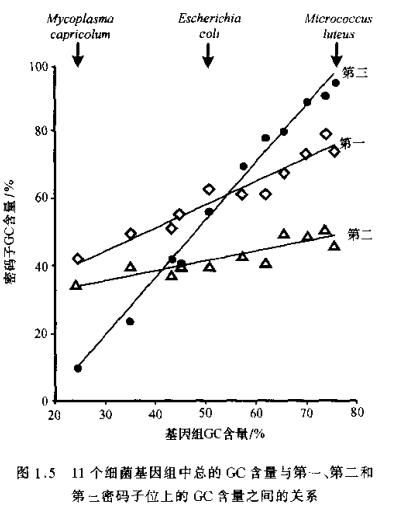
因此,密码子的使用受到突变压和净化选择的双重控制。